
Die Familie der ADAMs (a disintegrin and metalloproteinases) umfaßt eine Gruppe membranständiger Typ I Glykoproteine (über 40 Vertreter), die sowohl in physiologische (Fertilisation, Myoblastenfusion, Neurogenese) als auch pathologische Prozesse (Arteriosklerose, Osteoarthritis, Alzheimer, Tumorgenese) involviert sind. Sie werden auch als MDCs bezeichnet und sind somit als Proteine mit einer Metalloproteinase-, Disintegrin-, und Cystein-reichen-Domäne klassifiziert. ADAMs stehen in enger Homologie zu den in Schlangengiften gefundenen SVMPs (snake venom metalloproteinases) der Klasse P-III. ADAMs und SVMPs gehören beide zur Familie der Adamalysine und diese wiederum zusammen mit den Familien der MMPs (matrix metalloproteinases), Astacinen (zuerst im Flußkrebs Astacus astacus beschrieben) und Serralysinen (bakterielle Proteinasen) zur Superfamilie der Metzinkine. Alle Vertreter der Superfamilie der Metzinkine sind zinkabhängige Proteinasen mit einem konservierten Methionin-Rest und einem Zinkbindungsmotiv HEXXHXXGXXH in der proteolytisch aktiven Domäne. Im allgemeinen sind ADAMs aus einem Signalpeptid, einer Pro-Domäne, einer Metalloproteinase-Domäne, einer Disintegrin-Domäne, einer Cystein-reichen Domäne, einer EGF(epidermal growth factor)-ähnlichen Domäne, einer Transmembranregion und einer zytoplasmatischen Domäne aufgebaut. Die Tabelle 1 gibt einen Überblick über die bis zum jetzigen Zeitpunkt klonierten und charakterisierten Vertreter dieser Familie. Die ebenfalls aufgeführten ADAMTSs (a disintegrin and metalloproteinase with thrombospondin type-1 modules), die anstelle einer Transmembran- und zytoplasmatischen Domäne mehrere TS(Thrombo-spondin)-Motive enthalten, stellen lösliche Vertreter der Familie der ADAMs dar.
ADAM | Alternative Bezeichnung | Charakteristika | Literatur | Organismus | Accession Number |
ADAM 1 | PH-30 a | involviert bei der Fusion von Spermatozoon und Oozyte; Pseudogen bei Primaten und Menschen | Wolfsberg et al. (1993), Perry et al. (1995), Jury et al. (1997), Jury et al. (1998) |
Bos taurus
|
AF086807
|
ADAM 2 | PH-30 b | Interaktion mit dem Oozyten-Integrin a6b1 | Almeida et al. (1995), Cho et al. (1998), Chen et al. (1999a), Chen et al. (1999b), Chen et al. (1999c) |
Bos taurus
|
AF086808
|
ADAM 3A | Cyritestin 1 bzw. 2 | ADAM 3A ist involviert bei der Adhäsion und Fusion von Spermatozoon und Oozyte; Pseudogen beim Menschen; ADAM 3B existiert in einer membrangebundenen und löslichen Form | Linder et al. (1997), Yuan et al. (1997), Adham et al. (1998), Frayne und Hall (1998) |
Homo sapiens | |
ADAM 4 | tMDC V | vorwiegend Testis-spezifische Expression | Wolfsberg et al. (1995) |
Mus musculus
| |
ADAM 5 | tMDC II | vorwiegend Testis-spezifische Expression; Pseudogen beim Menschen | Wolfsberg et al. (1995), Frayne et al. (1999) |
Cavia porcellus
| |
ADAM 6 | tMDC IV | vorwiegend Testis-spezifische Expression | Wolfsberg et al. (1995) |
Cavia porcellus
| |
ADAM 7 | EAP I | Funktion bei der Spermienreifung | Perry et al. (1995), Cornwall und Hsia (1997) |
Homo sapiens
| |
ADAM 8 | MS2 | Funktion bei der Extravasation von Leukozyten und der Differenzierung von Osteoclasten | Yashida et al. (1990), Yoshiyama et al. (1997), Choi et al. (2001) |
Homo sapiens
| |
ADAM 9 | MDC 9 | wird in vielen Geweben exprimiert; a-Sekretaseaktivität; spaltet die Insulin-B-Kette, Gelatin, Fibronektin und b-Casein; Interaktion mit SH3 Proteinen; Shedding von HB-EGF; Interaktion mit den Integrinen a6b1 und avb5 | Izumi et al. (1998), Howard et al. (1999), Koike et al. (1999), Roghani et al. (1999), Weskamp et al. (1999), Nath et al. (2000), Schwettmann und Tschesche (2001), Zhou et al. (2001) |
Bos taurus:
| |
ADAM 10 | MADM | wird in vielen Geweben exprimiert; notwendig für die Axon-Extension in Drosophila melanogaster; a-Sekretaseaktivität; prozessiert NOTCH; spaltet proTNF-a, MBP und Typ IV Kollagen | Fambrough et al. (1996), Howard et al. (1996), Lunn et al. (1997), Pan und Rubin (1997), Rosendahl et al. (1997), Sotillos et al. (1997), Chubinskaya et al. (1998), Glynn et al. (1998), Dallas et al. (1999) |
Bos taurus
|
Z21961
|
ADAM 11 | MDC | Tumorsuppressorgen | Emi et al. (1993) |
Homo sapiens | |
ADAM 12 | Meltrin a | spaltet a-Makroglobulin; IGFBP-3 und -5; bindet an a-Actinin-2; beteiligt an der Myoblastenfusion; wird inhibiert durch TIMP 3 | Yagami-Hiromasa et al. (1995), Galliano et al. (2000), Loechel et al. (2000), Shi et al. (2000) |
Homo sapiens | |
ADAM 13 | Interaktion der zytoplasmatischen Domäne mit X-PACSIN2 in vivo | Alfandari et al. (1997), Cousin et al. (2000) | Xenopus laevis | ||
ADAM 14 | adm-1 | mögliche Funktion bei der Embryonalentwicklung in Caenorhabditis elegans | Podbilewicz (1996) | Caenorhabditis elegans | |
ADAM 15 | Metargidin | enthält ein RGD-Motiv; Interaktion mit den Integrinen avb3, a5b1 und a9b1 | Krätzschmar et al. (1996), Lum et al. (1998), Zhang et al. (1998) |
Homo sapiens | |
ADAM 16 | MDC 16 | möglicherweise involviert bei der Fertilisation von Xenopus laevis | Shilling et al. (1997) | Xenopus laevis | |
ADAM 17 | TACE | a-Sekretaseaktivität; prozessiert proTNF-a, TNF-a-Rezeptor, NOTCH 1-Rezeptor, L-Selektin, TRANCE, HERJM-a; wird inhibiert durch TIMP 3; Aktivierung durch NO möglich | Black et al. (1997), Moss et al. (1997), Amour et al. (1998), Peschon et al. (1998), Lum et al. (1999), Brou et al. (2000), Rio et al. (2000), Zhang et al. (2000), Lee et al. (2001) |
Homo sapiens | |
ADAM 18 | tMDC III | Spermienoberflächenprotein | Frayne et al. (1998) | Macaca fascicularis | |
ADAM 19 | Meltrin b | möglicherweise eine Funktion bei der Differenzierung von Osteoblasten; spaltet a-Makroglobulin | Inoue et al. (1998), Fritsche et al. (2000), Wei et al. (2001) | Mus musculus | |
ADAM 20 | Testis-spezifische Expression | Poindexter et al. (1999), van Huijsduijnen (1998) | Homo sapiens | ||
ADAM 21 | Testis-spezifische Expression | Poindexter et al. (1999), van Huijsduijnen (1998) | Homo sapiens | ||
ADAM 22 | MDC 2 | vorwiegend im Gehirn exprimiert | Sagane et al. (1998), Poindexter et al. (1999) | Homo sapiens | |
ADAM 23 | MDC 3 | vorwiegend im Gehirn exprimiert | Sagane et al. (1998), Poindexter et al. (1999) | Homo sapiens | |
ADAM 24 | Testase 1 | Testis-spezifische Expression | Zhu et al. (1999) | Mus musculus | |
ADAM 25 | Testase 2 | Testis-spezifische Expression | Zhu et al. (1999) | Mus musculus | |
ADAM 26 | Testase 3 | Testis-spezifische Expression | Zhu et al. (1999) | Mus musculus | |
ADAM 28 | MDC-Lm | prozessiert autokatalytisch die eigene Pro-Domäne; spaltet MBP | Roberts et al. (1999), Howard et al. (2000), Howard et al. (2001) |
Homo sapiens | |
ADAM 29 | Testis-spezifische Expression; drei Spleißvarianten | Cerretti et al. (1999a), Cerretti et al. (1999b) |
Homo sapiens | ||
ADAM 30 | Testis-spezifische Expression; zwei Spleißvarianten | Cerretti et al. (1999a), Cerretti et al. (1999b) |
Homo sapiens | ||
ADAMTS 1 | METH1 | spaltet Aggrecan zwischen Glu(1871) und Leu(1872); spaltet a-Makroglobulin | Kuno und Matsushima (1998), Kuno et al. (1999), Kuno et al. (2000), Shindo et al. (2000) | Mus musculus | |
ADAMTS 2 | PCNP | als Prokollagen-N-Protease beteiligt an der Kollagenbiosynthese | Colige et al. (1997) | Homo sapiens | |
ADAMTS 3 | Kloniert aus Hirngewebe | Nagase et al. (1997) | Homo sapiens | ||
ADAMTS 4 | Aggrecanase 1 | spaltet Aggrecan zwischen Glu(373) und Ala(374); spaltet Brevican zwischen Glu(395) und Ser(396); wird inhibiert durch TIMP 3; Transkription wird durch Ab stimuliert | Tortorella et al. (1999), Matthews et al. (2000), Satoh et al. (2000), Nakamura et al. (2000), Kashiwagi et al. (2001) |
Homo sapiens | |
ADAMTS 5 | Aggrecanase-2 | spaltet Aggrecan zwischen Glu(373) und Ala(374); wird inhibiert durch TIMP 3 | Abbaszade et al. (1999), Kashiwagi et al. (2001) |
Homo sapiens | |
ADAMTS 6 | vorwiegend in der Plazenta exprimiert | Hurskainen et al. (1999) | Homo sapiens | ||
ADAMTS 7 | wird in vielen Geweben exprimiert | Hurskainen et al. (1999) | Homo sapiens | ||
ADAMTS 8 | stört die Angiogenese in vivo | Vasquez et al. (1999) | Homo sapiens | ||
ADAMTS 9 | wird in allen bisher getesteten fötalen Geweben exprimiert | Clark et al. (2000) | Homo sapiens | ||
ADAMTS 12 | signifikante Expression nur in fötalen Lungen; spaltet a-Makroglobulin | Cal et al. (2001) | Homo sapiens |
Die Familie der ADAMs, deren erste Vertreter Ende der 80er Jahre entdeckt wurden, weisen Strukturhomologien zu den in Schlangengiften gefundenen SVMPs der Klasse P-III auf. Aus diesem Grund wurde vermutet, daß ADAMs ähnliche Eigenschaften wie SVMPs besitzen, also möglicherweise proteolytisch aktiv oder potentielle Integrinliganden seien. Die SVMPs sind lösliche Bestandteile von Schlangenseren und werden als Proenzyme in den Giftdrüsen exprimiert und anschließend in ihre aktive Form übergeführt (Jia et al., 1996). Sie sind für die hämorrhagische Aktivität der Schlangengifte verantwortlich, welche unter anderem zu einer Degradation der Membranproteine der Blutgefäßwand führen (Markland, 1997). Eine Vielzahl dieser Schlangengiftkomponenten beeinflußt das Blutgerinnungssystem des Menschen und anderer Säuger (Markland, 1997). Einige von ihnen führen z.B. zur Degradation des Fibrinogens oder Fibrins aber auch zur Aktivierung des Plasminogens oder des Prothrombins. Auch die Gerinnungsfaktoren V oder X können von Schlangengiften aktiviert werden. Des weiteren hemmen oder aktivieren Schlangengifte die Aggregation der Thrombozyten. Der wesentliche Unterschied zwischen ADAMs und SVMPs besteht darin, daß SVMPs lösliche Proteine mit proteolytischer Aktivität sind, wohingegen nur 70% der zur Zeit bekannten ADAMs ein potentielles Zinkbindungsmotiv aufweisen. Die bisher bekannten Vertreter der ADAMs beinhalten 800 bis 1200 Aminosäuren und sind, mit Ausnahme von Spleißvarianten und den ADAMTSs, membrangebundene Proteine die in einer Vielzahl von unterschiedlichen Spezies (Mensch, Primaten, Nagern, Amphibien, Würmern und Hefen) entdeckt worden sind. Ein Vergleich der Domänenstruktur von ADAMs, ADAMTSs, SVMPs und dem einfachsten Vertreter der MMPs, dem Matrilysin (MMP 7), ist in der folgenden Abbildung gegeben:
Auffällig sind vor allem die TS-Motive innerhalb der ADAMTSs. Es sind Wiederholungen mehrerer TS1-Module, die normalerweise innerhalb multifunktioneller Proteine der extrazellulären Matrix, die Adhäsion, Motilität und das Wachstum von Zellen beeinflussen. Über die Funktion der innerhalb der Familie der ADAMTSs nur wenig konservierten Spacer-Regionen ist bisher nur wenig bekannt. Allen in der Abbildung dargestellten Proteinen ist ein Signalpeptid am N-Terminus gemeinsam, welches die Proteine in den sekretorischen Pfad durch das endoplasmatische Reticulum und den Golgi-Apparat dirigiert. Es wird kurz vor der Sekretion bzw. dem Einbau in die Plasmamembran entfernt. Die Funktionen der weiteren in der Abbildung dargestellten Domänen sollen im folgenden kurz besprochen werden.
Die Pro-Domäne beinhaltet im allgemeinen 200 Aminosäurereste und wird in vielen Fällen durch eine Furin-Spaltstelle RX(K/R)R von der Metalloproteinase-Domäne getrennt (Hurskainen et al., 1999). Furin oder furinartige Proteinasen des Subtilisin-Typs spalten im Golgi-Apparat hinter dieser Erkennungssequenz und prozessieren somit die Proform des Moleküls, wie z.B. im Fall von ADAM 9, 12, 15 und ADAMTS 1 (Lum et al., 1998; Kuno et al., 1999; Loechel et al., 1999; Roghani et al., 1999). Dieser Vorgang ist nicht von einer proteolytischen Aktivität der Metalloproteinase-Domäne abhängig. Mutationsexperimente, bei denen der Glutamat-Rest im katalytischen Zentrum gegen Glutamin oder Alanin ausgetauscht worden ist, belegen, daß die Prozessierung trotzdem vonstatten geht. ADAM 28 ist in diesem Zusammenhang die erste bekannte Ausnahme, denn Untersuchungen zeigten, daß die Entfernung der Pro-Domäne zumindest partiell autoproteolytisch, also in Abhängigkeit von der proteolytisch aktiven Metalloproteinase-Domäne verläuft (Howard et al., 2000). Die Pro-Domäne der ADAMs beinhaltet in vielen Fällen das von den MMPs und anderen Reprolysinen bekannte cysteine-switch-Motiv. Es besteht aus einem in der Pro-Domäne konservierten Cystein-Rest, über dessen freie Thiolgruppe das im katalytischen Zentrum komplexierte Zink(II)ion zusätzlich koordiniert wird. Diese Komplexierung des katalytisch relevanten Zink(II)ions gewährleistet die Latenz der Proenzyme, und erst durch die Aufhebung der Wechselwirkung der freien Thiolgruppe des Cysteins mit diesem Zink(II)ion (bespielsweise durch Oxidation, Methylierung oder auch durch Interaktion mit Übergangsmetallen wie Quecksilber(II)ionen) werden die Enzyme aktiviert (van Wart und Birkedal-Hansen, 1990; Grams et al., 1993). Die Regulation der proteolytischen Aktivität während der Biosynthese ist notwendig um eine vorzeitige und unkontrollierte Aktivierung dieser Proteinasen zu verhindern. Belege dafür, daß dieses Modell auch auf die Familie der ADAMs übertragbar ist, liefern Studien an ADAM 12 und 17, die zeigen, daß die Pro-Domäne die Aktivität der Metalloproteinase-Domäne inhibiert und darüber hinaus auch die korrekte Konformation des Moleküls bedingt (Loechel et al., 1999; Milla et al., 1999).
Die Metalloproteinase-Domäne der ADAMs besteht aus bis zu 200 Aminosäuren und gehört zu der Superfamilie der Metzinkine (Bode et al., 1993). Strukturell verfügen alle Metzinkine über ein fünfsträngiges b-Faltblatt und drei a-Helices in ihrer katalytisch aktiven Domäne. In dieser Domäne wird ein Zink(II)ion von drei Histidin-Resten im katalytischen Zentrum koordiniert, welche das für diese Familie typische Zinkbindungsmotiv HEXXHXXGXXH generieren. Die vierte Koordinationsstelle des Zink(II)ions wird von einem Wassermolekül eingenommen, so daß sich insgesamt eine tetraedrische Koordinationssphäre für das Zink(II)ion ergibt. Zusätzlich zu diesem katalytisch relevanten Zink(II)ion befinden sich auch noch ein strukturstabilisierendes Zink(II)ion und zwei bis drei strukturstabilisierende Calcium(II)ionen innerhalb dieser Domäne (Bode et al., 1999). Der Glutamat-Rest innerhalb des Zinkbindungsmotives fungiert als katalytische Base und der Glycin-Rest ermöglicht einen Umkehrpunkt in der Struktur (Jiang und Bond, 1992). Der in der Superfamilie der Metzinkine konservierte Methionin-Rest befindet sich in den meisten Fällen 20 - 30 Aminosäuren C-terminal vom Zinkbindungsmotiv und trägt wahrscheinlich zur Stabilisierung der proteolytisch aktiven Struktur bei.
Neben diesen Gemeinsamkeiten zwischen ADAMs und den übrigen Vertretern der Metzinkine existieren zwei grundlegende Unterschiede. Zum einen fehlt in fast 30% aller bisher bekannten ADAMs das funktionelle Zinkbindungsmotiv, dazu gehören die ADAMs 2 -7, 11, 14, 18, 22, 23 und 29, so daß anzunehmen ist, daß diese ADAMs keinerlei proteolytische Funktion ausüben. Zum anderen ist die Metalloproteinase-Domäne in einigen maturierten Formen der ADAMs nicht mehr existent, dazu gehören z.B. ADAM 1 und 2 (Fertilin a und b) (Blobel et al., 1992). Die Frage nach der Funktion und dem Verbleib nach Prozessierung der Metalloproteinase-Domäne in diesen beiden Proteinen ist bis heute unbeantwortet geblieben (Blobel, 2000). Im überwiegenden Teil (70%) der bis heute bekannten ADAMs ist das funktionelle Zinkbindungsmotiv der Metzinkine vorhanden. Das sind die ADAMs 1, 8 - 10, 12, 13, 15 - 17, 19 - 21, 24 - 26, 28, 30, 31 und die ADAMTSs 1 - 9 bzw. 12. Es ist wahrscheinlich, daß diese Proteine - oder eine ihrer prozessierten Formen - proteolytische Aktivität besitzen. Für einen Großteil dieser ADAMs und ADAMTSs steht die biochemische Charakterisierung noch aus und somit auch die Beantwortung der Frage nach ihren natürlichen Substraten und Inhibitoren.
Nur für eine kleine Gruppe proteolytisch aktiver ADAMs und ADAMTSs sind bisher in vivo-Substrate identifiziert worden. In vielen Fällen handelt es sich bei diesen Substraten um zelloberflächenassoziierte Proteine, die durch die proteolytische Prozessierung in einen löslichen Teil und einen weiterhin membrangebundenen Teil gespalten werden. Dieser Vorgang, das sogenannte shedding (shed = verlieren, abwerfen) führte zu einer alternativen Bezeichnung der ADAMs, die danach auch als sheddases bezeichnet werden. Beispielsweise ist ADAM 9 zusammen mit dem Isotyp d der Proteinkinase C (PKCd) für das TPA (12-O-Tetradecanoylphorbol-13-acetat) stimulierte shedding von proHB-EGF (heparin-binding EGF-like growth factor) verantwortlich, wobei eine direkte Interaktion von PKCd; und der zytoplasmatischen Domäne von ADAM 9 vermutet wird (Izumi et al., 1998). Die Prozessierung des zelloberflächenassoziierten Rezeptors NOTCH durch ADAM 10 (KUZ), bei der NOTCH in einen extrazellulären und intrazellulären Teil gespalten wird, ist ein wichtiger Schritt bei der lateralen Inhibierung der Neurogenese bei Drosophila melanogaster (Pan und Rubin, 1997; Sotillos et al., 1997). Bei der Ratte scheint ADAM 10 eine Funktion bei der Aktivierung von Homologen des NOTCH-Rezeptors zu haben, wobei diese auf den Knochen beschränkt ist und keinen Einfluß z.B. auf die Entwicklung des Knorpels zu haben scheint (Dallas et al., 1999). Das wohl bekannteste Beispiel ist die Prozessierung der membrangebundenen Proform des Zytokins TNF(tumor necrosis factor)-a, welches in Entzündungsprozesse involviert ist und dessen lösliche und somit aktive Form durch ADAM 17 (TACE (tumor necrosis factor-a-converting enzyme)) generiert wird (Bazzoni und Beutler, 1996; Black et al., 1997; Moss et al., 1997). Bemerkenswert sind in diesem Zusammenhang Hinweise darauf, daß die Prozessierung von proTNF-a; nicht nur auf der Zelloberfläche sondern möglicherweise schon in einem intrazellulären Kompartiment vonstatten geht. Unterstützt wird diese Vermutung durch die Beobachtungen, daß sowohl ADAM 17 als auch proTNF-a bevorzugt in einem perinukleären Zellkompartiment lokalisiert vorliegen (Schlöndorff et al., 2000), und daß durch die Stimulation von Zellen, z.B. mit PMA (Phorbol-12-myristat-13-acetat), ein Reagenz welches normalerweise die shedding-Aktivität erhöht, ein verminderte Präsenz des auf der Zelloberfläche lokalisierten ADAM 17 zu beobachten ist. Möglicherweise wird ADAM 17 nach Stimulation mit PMA internalisiert und somit von der Zelloberfläche entfernt. (Doedens und Black, 2000). Ein Knockout der ADAM 17-Expression in Mäusen geht mit einer abnormen Entwicklung der Lungen einher, ein normalerweise äußerst Zytokin- und Wachstumsfaktor-abhängiger Prozeß. Weiterführende Untersuchungen zeigten, daß diese Entwicklungsdefekte nicht auf eine mangelnde Prozessierung von proTNF-a; zurückzuführen sind, so daß die sheddase-Aktivität von ADAM 17 nicht nur auf proTNF-a; beschränkt zu sein scheint (Zhao et al., 2001). ADAMTS 2 prozessiert Prokollagen I und II, ein essentieller Schritt bei der Ausbildung von Kollagenfasern. Ein Ausschalten der ADAMTS 2-Expression in vivo führt zu einer erhöhten Fragilität des Bindegewebes, welche durch die Akkumulation von Prokollagen und der Ausbildung abnormer Kollagenfasern hervorgerufen wird (Colige et al., 1997; Li et al., 2001, Tang, 2001). Die Prozessierung von Aggrecan (ein Proteoglykan und Hauptbestandteil des Knorpels) durch ADAMTS 4 und 5 ist für die Elastizität und Festigkeit des Knorpelgewebes entscheidend (Tortorella et al., 1999).
Für einige Vertreter der ADAMs war es möglich in vitro Substrate zu identifizieren. ADAM 9 spaltet beispielsweise die Insulin-B-Kette und mehrere synthetische Peptide (Roghani et al., 1999). Die Stimulation mit PMA führt dazu, daß ADAM 9 eine a-Sekretaseaktivität gegenüber coexprimierten APP(amyloid precursor protein)-Analoga in COS-7 Zellen aufweist (Koike et al., 1999). Durch Zymographie konnten Gelatin, b-Casein und Fibronektin in vitro als Substrate identifiziert werden (Schwettmann und Tschesche, 2001). ADAM 10 spaltet Typ IV Kollagen in vitro und prozessiert ein synthetisches Peptid, welches der proTNF-a-Spaltstelle nachempfunden wurde. (Lunn et al., 1997; Rosendahl et al., 1997; Millichip et al., 1998). Im Gegensatz dazu weisen neuere Untersuchungen unter Verwendung von Antisense-Oligonukleotiden daraufhin, daß die Prozessierung von proTNF-a durch ADAM 10 nicht in den Leukozytenzellinien Jurkat und THP-1 gefunden wird (Condon et al., 2001). ADAM 12 spaltet a2-Makroglobulin, prozessiert IGFBP(insulin-like growth factor-binding protein)-3 und IGFBP-5 und wird durch TIMP(tissue inhibitor of metalloproteinases) 3 inhibiert (Loechel et al., 1998, Loechel et al. 2000). ADAM 17 prozessiert den extrazellulären Teil des NOTCH 1-Rezeptors (Brou et al., 2000). Das APP wird durch die potentiellen a-Sekretasen ADAM 9, 10 und 17 gespalten (Buxbaum et al., 1998; Lammich et al., 1999; Schlöndorff und Blobel, 1999). ADAM 19 führt zur Bildung eines kovalenten Komplexes mit und zur Spaltung von a-Makroglobulin (Wei et al., 2001). Hinweise auf die proteolytische Aktivität von ADAM 28 gibt die Beobachtung, daß die Abspaltung der Pro-Domäne - zumindest teilweise - autoproteolytisch verläuft (Howard et al., 2000) und erst kürzlich konnte gezeigt werden, daß ADAM 28 das MBP (myelin basic protein) spaltet (Howard et al., 2001). ADAMTS 1 prozessiert Aggrecan und bildet einen kovalenten Komplex mit a-Makroglobulin (Kuno et al., 1999; Kuno et al., 2000). Für ADAMTS 4 konnte Brevican, ein vorwiegend im ZNS (Zentralnervensystem) exprimiertes Proteoglykan als in vitro-Substrat identifiziert werden (Nakamura et al., 2000). ADAMTS 4 ist ebenso wie ADAMTS 5 durch die N-terminale inhibitorische Domäne von TIMP 3 hemmbar (Kashiwagi et al., 2001).
Die einzige bisher bekannte räumliche Struktur einer ADAM-Metalloproteinase-Domäne ist die von ADAM 17, welches zur Analyse der Struktur mit einem Hydroxamatinhibitor cokristallisiert wurde. Vergleicht man die Struktur von ADAM 17 mit denen von MMPs, so finden sich neben Gemeinsamkeiten im räumlichen Aufbau der Substratbindungsstelle auch einige Unterschiede. Die S3'-Spezifitätstasche von ADAM 17 ist im Vergleich zu der von MMPs wesentlich tiefer ausgebildet und weist eine Verbindung zur S1'-Spezifitätstasche auf. Darüber hinaus fehlt in der Metalloproteinase-Domäne von ADAM 17 eine der für die Adamalysine typische Calcium-Bindungsstelle (siehe 2.2.2) (Maskos et al., 1998). Die strukturellen Gemeinsamkeiten und Unterschiede zwischen ADAM 17 und den MMPs könnten der Grund für die Beobachtung sein, daß zwar TIMP 3 die Metalloproteinase-Domäne von ADAM 17 zu hemmen vermag (Amour et al. 1998; Lee et al., 2001), TIMP 1, 2 und 4 dazu aber nicht in der Lage sind. ADAM 10 zeigt ein ähnliches Inhibierbarkeitsprofil, da es von TIMP 1 und 3 aber nicht von TIMP 2 und 4 gehemmt wird (Amour et al., 2000). Bei diesen Untersuchungen wurde jedoch der Einfluß der Disintegrin-, Cystein-reichen und EGF-ähnlichen Domäne der betreffenden ADAMs außerachtgelassen, da stets nur die Metalloproteinase-Domänen alleine untersucht wurden. Möglicherweise zeigen die Gesamtmoleküle ein anderes Verhalten gegenüber Inhibitoren und Substraten.
Die Disintegrin-Domäne der ADAMs zeigt sehr hohe Sequenzhomologien zu den SVMPs der Klasse P-III, besteht aus 60 bis 90 Aminosäuren und beinhaltet 6 bis 15 Cystein-Reste (Bjarnason und Fox, 1995). Die Bindung an Integrine wird durch einen 13 bis 14 Aminosäuren umfassenden loop vermittelt, an dessen Spitze eine für das jeweilige Disintegrin charakteristische Tripeptidsequenz lokalisiert ist. Diese Tripeptidsequenz und die sie N- und C-terminal flankierenden Aminosäuren bestimmen die Spezifität und Affinität gegenüber Integrinen. Die Disintegrin-Domänen der SVMPs der Klasse P-III enthalten in den meisten Fällen ein RGD-Tripeptid, wodurch sie beispielsweise mit dem Plättchenintegrin aIIb3 interagieren und somit die Aggregation der Thrombozyten verhindern. Man vermutet, daß der loop innerhalb der Disintegrin-Domäne in die ,Tasche" eines Integrin-Heterodimers hineinragt und dadurch die Wechselwirkung zwischen Disintegrin und Integrin initiiert (Wolfsberg und White, 1996). Ein wesentlicher Unterschied zwischen den Disintegrin-Domänen der ADAMs und SVMPs ist, daß diese Domänen bei den membranverankerten ADAMs eher Zell-Zell-Interaktionen vermitteln, wohingegen sie bei den löslichen SVMPs der Klasse P-III derartige Interaktionen bevorzugt unterbinden bzw. lösen.
In mehreren in vitro-Studien konnten für die Disintegrin-Domänen einiger ADAMs Interaktionspartner aus der Familie der Integrine identifiziert werden. ADAM 2 zeigt bei der Fertilisation eine Wechselwirkung mit dem Oozyten-Integrin a6b1 (Chen et al., 1999a; Chen et al., 1999b). Diese Interaktion wird durch das Tripeptid ECD im Disintegrin-Loop hervorgerufen und der darin enthaltene Aspartat-Rest ist maßgeblich an dieser Wechselwirkung zwischen Disintegrin und Integrin beteiligt (Bigler et al., 2000; Zhu et al., 2000). Ein lösliches Tripeptid und ein dagegen gerichteter Antikörper, nachempfunden dem Tripeptid-Motiv in der Disintegrin-Domäne von ADAM 3, vermögen beide die Adhäsion und Fusion von Spermatozoon und Oozyte zu inhibieren (Yuan et al., 1997). Weitergehende Untersuchungen konnten zeigen, daß hierbei auch der Glutamin-Rest an Position 7 des Disintegrin-loops maßgeblich beteiligt ist (Takahashi et al., 2001). Die Interaktion der Disintegrin-Domäne von ADAM 9 mit dem Integrin a6b1 auf der Oberfläche der Fibroblastenzellinie HT1080 führt zu einer abgerundeten Morphologie und gesteigerter Motilität dieser Zellen, im Vergleich zu ihrer Motilität auf Laminin (Nath et al., 2000). Des weiteren zeigt ADAM 9 eine RGD-unabhängige Interaktion mit dem Integrin vb5 auf Myelomzellen (Zhou et al., 2001). Auch für ADAM 12 und 15 konnte eine RGD-unabhängige Interaktion mit dem Integrin a9b1 gezeigt werden, die Zell-Zell-Interaktionen unterstützt (Eto et al., 2000). Humanes ADAM 15 interagiert aber auch in RGD-abhängiger Weise mit den Integrinen avb3 und a5b1 wodurch die Ausbildung von Zell-Zell-Kontakten induzieren werden können (Zhang et al., 1998; Krätzschmar et al., 1996). ADAM 23 interagiert mit dem Integrin avb3 über einen RGD-unabhängigen Mechanismus (Cal et al., 2000).
Die Funktionen dieser beiden Domänen sind bis heute noch nicht gut verstanden. Die Cystein-reiche Domäne besteht aus über 160 Aminosäuren mit 10 bis 14 Cystein-Resten. Einige Cystein-reiche Domänen besitzen in einem 23 Aminosäuren umfassenden a-helicalen Bereich eine Strukturhomologie zu viralen Fusionsproteinen. Zu ihnen gehören ADAM 1, 9, 11 und 12 und man vermutet, daß diese ADAMs eine Funktion bei Zell-Zell-Fusionsprozessen haben (Wolfsberg und White, 1998). Die EGF-ähnliche Domäne umfaßt ungefähr 40 Aminosäuren mit 6 Cysteinen. Möglicherweise sind diese beiden Domänen für die Interaktion der ADAMs mit anderen Proteinen verantwortlich, wie z.B. Chaperonen bei der Biosynthese oder Interaktionspartnern und Substraten auf der Zelloberfläche. Diese Wechselwirkungen führen die ADAMs möglicherweise an ihren physiologischen Wirkort und regulieren darüber hinaus auch ihre Aktivität. Unterstützt werden diese Hypothesen durch die Beobachtungen, daß die Cystein-reiche Domäne von ADAM 17 an der Freisetzung der Pro-Domäne und beim shedding des Interleukin 1 Typ II-Rezeptors beteiligt ist (Milla et al., 1999; Reddy et al., 2000). Die lösliche Spleißvariante von ADAM 12, das sogenannte ADAM 12 S, bindet an das IGFBP-3 über seine Cystein-reiche Domäne und prozessiert dieses dann proteolytisch (Shi et al., 2000). Des weiteren unterstützt die Cystein-reiche Domäne von ADAM 12 die Adhäsion von Tumorzellen, z.B. der Mammakarzinomzellinie MDA-MB-231 (Iba et al., 1999).
ADAMs sind, mit Ausnahme von bestimmten Spleißvarianten und den ADAMTSs membranständige Proteine des Typs I und enthalten somit in der Nähe ihres C-Terminus eine hydrophobe Region mit der sie in der Plasmamembran verankert sind. Die Spleißvarianten von ADAM 11, 12, 17 und 28 bringen jedoch auch lösliche Formen dieser Proteine hervor, denen die Transmembran-Region und die zytoplasmatische Domäne fehlen (Emi et al., 1993; Gilpin et al., 1998; Cerretti et al., 1999; Roberts et al., 1999). Ob alle bisher klonierten ADAMs sowohl in membranverankerten als auch löslichen Formen existieren - entweder durch alternatives Spleißen oder shedding-Prozesse generiert - ist bis jetzt noch unbekannt.
Die zytoplasmatische Domäne der ADAMs variiert in ihrer Länge von 40 bis 250 Aminosäuren und ist möglicherweise an intrazellulären Signaltransduktionsprozessen beteiligt. Bei einigen ADAMs weist sie Prolin-reiche Sequenzabschnitte auf, die auf eine Anbindung an das Zyto-skelett oder auf eine Funktion als SH(src homology)3-Domänen-Ligand hindeuten. ADAM 9 und 15 interagieren z.B. mit den SH3-Domänen-tragenden Proteinen Endophilin I und SH3PX1 (Howard et al., 1999), wobei eine Auswirkung auf die intrazelluläre Prozessierung, den Transport oder die subzelluläre Lokalisation dieser beiden ADAMs vermutet wird. Eine Interaktion der zytoplasmatischen Domäne von ADAM 12 konnte mit der Protein-Tyrosin-Kinase Src, ein ebenfalls SH3-Domänen-tragendes Protein beobachtet werden, wobei es zu einer Aktivierung der Kinase durch Autophosphorylierung kommt (Kang et al., 2000). Des weiteren tritt ADAM 12 über seine zytoplasmatische Domäne mit dem muskelspezifischen Aktin-Bindungsprotein a-Aktin-2 in Wechselwirkung, ein Vorgang, der für die Myoblastenfusion essentiell ist (Galliano et al., 2000). Ein weiteres Charakteristikum der zytoplasmatischen aber auch der Transmembran-Domäne von ADAM 12 ist eine Signalsequenz, die eine verlangsamte Prozessierung im trans-Golgi-Netzwerk bewirkt (Hougaard et al., 2000). Möglicherweise ist bei ADAM 15 eine derartige Signalsequenz in diesen beiden Domänen ebenfalls präsent, da ADAM 15 größtenteils in einem intrazellulären Kompartiment anstatt auf der Zelloberfläche gefunden wurde (Lum et al., 1998). Auch für ADAM 13 aus Xenopus laevis konnte eine Interaktion mit SH3-Domänen-tragenden Proteinen gezeigt werden (Cousin et al., 2000). Die zytoplasmatische Domäne von ADAM 17 interagiert mit einem bei der Mitose involvierten Protein, dem sogenannten MAD(mitotic arrest deficient)2, wobei ein weiterer Vertreter dieser Familie, das MAD2b; eine Interaktion mit der zytoplasmatischen Domäne von ADAM 9 zeigt (Nelson et al., 1999).
Die löslichen ADAMTSs enthalten zwei TS-Motive, die in sich aus mehreren TS1-Modulen aufgebaut sein können. Das erste, näher am N-Terminus liegende TS-Motiv befindet sich zwischen der Disintegrin- und Cystein-reichen Domäne. Das zweite TS-Motiv wird durch eine bisher nicht näher charakterisierte Spacer-Region von der Cystein-reichen Domäne getrennt. Bei einigen ADAMTSs folgt auf das zweite TS-Motiv nochmals eine Spacer-Region (Abb. 2.1). Die TS-Motive beeinflussen zusammen mit den Spacer-Regionen die Interaktion der ADAMTSs mit ihrer Umgebung. Beispielsweise zeigen die TS-Motive von ADAMTS 1 eine ausgeprägte Affinität zu Heparin oder sind zusammen mit der Spacer-Region für eine starke Wechselwirkung mit der extrazellulären Matrix verantwortlich (Kuno et al., 1997; Kuno und Matsushima, 1998). An der Spaltung von Aggrecan durch ADAMTS 1 ist die Spacer-Region ebenfalls beteiligt (Kuno et al., 2000). Auch bei ADAMTS 4 ist das TS-Motiv entscheidend an der Erkennung und Spaltung von Aggrecan beteiligt (Tortorella et al., 2000). Das kürzlich beschriebene ADAMTS 12 enthält acht TS1-Module, die als ein-, drei- und viergliederige Modulgruppierung die drei TS-Motive des Moleküls aufbauen. Eine weitere Besonderheit ist die intrazelluläre Prozessierung von ADAMTS 12 in ein 120 kDa N-terminales, proteolytisch aktives (spaltet a-Makroglobulin in vitro) und ein 83 kDa C-terminales Fragment (Cal et al., 2001).
Das humane ADAM 15 (hADAM 15, hMDC 15) oder auch Metargidin ist ein 814 Aminosäuren umfassendes Protein, dessen cDNA erstmals von Krätzschmar und Kollegen (1996) beschrieben worden ist. Durch RT-PCR konnte die cDNA aus der humanen Mammakarzinomzellinie MDA-MB-468 kloniert werden. Anhand von Northern-Blot-Analysen konnte gezeigt werden, daß das Primärtranskript in vielen Geweben, wie z.B. Herz, Niere, Lunge oder auch im Skelettmuskel zu finden ist. Die alternative Bezeichnung Metargidin soll die Zugehörigkeit zu den METAlloproteinasen mit einem RGD-Motiv innerhalb der DisintegrIN-Domäne symbolisieren (META RGiD IN). Dieses RGD-Motiv im hADAM 15 ist einzigartig unter allen bisher bekannten ADAMs. Die Maus- und Ratten-Homologe des ADAM 15 enthalten die Tripeptidsequenz TDD (Lum et al., 1998; Bosse et al., 2000). Abbildung 2 gibt einen Überblick über die Domänenstruktur des hADAM 15 mit seinen wichtigsten Charakteristika (rot gekennzeichnet): Modifiziertes cysteine-switch-Motiv GHTCALS innerhalb der Pro-Domäne; Furin-Spaltstelle RRRR zwischen Pro-und Metalloproteinase-Domäne; Zinkbindungsmotiv HELGHSLGLDH innerhalb der Metalloproteinase-Domäne, RGD-Tripeptid RGDC innerhalb der Disintegrin-Domäne, zwei Prolin-reiche Sequenzen PPPPRKP und RPAPPPP innerhalb der zytoplasmatischen Domäne.

Das Signalpeptid am N-Terminus dirigiert das ADAM 15 Molekül in den sekretorischen Pfad durch das endoplasmatische Reticulum und den Golgi-Apparat (Lum et al., 1998). Diesem Signalpeptid folgt die Pro-Domäne innerhalb der sich ein Cystein-Rest befindet, der für die Latenz der Metalloproteinase-Domäne über einen cysteine-switch-artigen Mechanismus verantwortlich sein könnte (siehe 2.2.1). Da aber das cysteine-switch-Motiv GHTCALS in hADAM 15 im Vergleich zum allgemeinen cysteine-switch-Konsensusmotiv PRCGVPD der Metzinkine modifiziert ist, kann die Regulation der proteolytischen Aktivität über diesen Mechanismus nicht uneingeschränkt angenommen werden (Birkedal-Hansen et al., 1993, McKie et al., 1997; Bode et al., 1999). Die vier Arginin-Reste zwischen der Pro- und der Metalloproteinase-Domäne sind charakteristisch für die Spaltstellen von Proprotein-Konvertasen, wie z.B. Furin. Die Aktivierung der Metalloproteinase-Domäne könnte also auch innerhalb des sekretorischen Pfades (s.o.), z.B. im Golgi-Apparat erfolgen. Für murines ADAM 15 konnte gezeigt werden, daß die Abspaltung der Pro-Domäne im trans-Golgi-Apparat erfolgt und Furin die Prozessierung des Proenzyms in vitro mimetisch vollzieht (Lum et al., 1998). Die Metalloproteinase-Domäne enthält ein für die Metzinkine beschriebenes Zinkbindungsmotiv mit drei Histidinen und einem katalytisch wirksamen Glutamat-Rest (HELGHSLGLDH), so daß eine proteolytische Aktivität des Moleküls diskutiert wird.
Die ersten Untersuchungen zur Affinität der Disintegrin-Domäne von hADAM 15 zu Integrinen wurden 1998 von Zhang und Kollegen durchgeführt. Sie fanden, daß die rekombinant als Fusionsprotein in E. coli hergestellte Disintegrin-Domäne von hADAM 15 mit dem Integrin avb3 interagiert. Eine Mutante bei der das RGD-Motiv gegen ein SGA-Motiv ausgetauscht worden war, zeigte diese Wechselwirkung nicht (Zhang et al., 1998). Die Autoren stellten zudem fest, daß die N-terminal flankierende Sequenz RPT vor dem RGD-Motiv die Affinität zum Liganden kontrolliert. Wurde das RPT-Motiv gegen ein NWK-Motiv ausgetauscht, zeigte die Disintegrin-Domäne eine zusätzliche Affinität zu dem Thrombozyten-Integrin aIIbb3. Weitere Hinweise für eine RGD-abhängige Interaktion der Disintegrin-Domäne mit potentiellen Integrin-Liganden kamen von Nath et al. (1999). Die Autoren verwendeten in ihrem Modell keine bakteriellen Expressionsprodukte, sondern stellten den gesamten extrazellulären Teil des Proteins (die sogenannte Ecto-Domäne) in COS-7 Zellen als Fc-Fusionsprotein rekombinant her. Der C-terminale Fc-Teil ermöglichte eine effektive Reinigung über Protein A-Sepharose und die Expression in eukaryontischen Zellen sollte gewährleisten, daß das Expressionsprodukt in der nativen Konformation mit einem korrekten Disulfidbrückenmuster gewonnen wurde. Die Autoren fanden eine Interaktion mit dem Integrin avb3, was die Ergebnisse von Zhang et al. (1998) bestätigte und zusätzlich eine Interaktion mit dem Integrin a5b1. Dabei war auffällig, daß nicht alle a5b1 exprimierenden Zellinien, die die Autoren verwendeten, diese Interaktion zeigten, sondern nur die T-Zellinie MOLT-4. Also scheint die Interaktion zwischen der Disintegrin-Domäne von hADAM 15 mit dem Integrin a5b1 nicht nur Integrin- sondern auch zellspezifisch zu sein. Die Disintegrin-Integrin-Wechselwirkung nahm mit ansteigender Temperatur zu und benötigte Calcium(II)- und Magnesium(II)ionen im umgebenden Medium. Durch die Gegenwart von Mangan(II)ionen wurde die Interaktion verstärkt (Nath et al., 1999). Die Interaktion mit dem Integrin avb3, welches essentiell an der Angiogenese beteiligt ist, mag im Hinblick auf die Tumorgenese von besonderem Interesse sein, da beobachtet worden ist, daß eine verstärkte Expression von avb3 auf Melanomzellen mit einer verstärkten Malignität einhergeht (Albeda et al., 1991; Montgomery et al., 1994). Darüber hinaus könnte, da hADAM 15 auf Endothelzellen gefunden worden ist (Herren et al., 1997), eine Interaktion von hADAM 15 mit avb3 die Adhäsion von Tumorzellen an das Endothel beeinflussen und somit einen Initiationsschritt bei der Neovaskularisierung darstellen (Nath et al., 1999). Das RGD-Motiv innerhalb der Disintegrin-Domäne von hADAM 15 weist jedoch noch eine Besonderheit auf. Direkt hinter dem Aspartat-Rest befindet sich ein Cystein-Rest RGDC, der ansonsten nur bei nicht-RGD-Typ-Disintegrinen gefunden wurde (Krätzschmar et al., 1996). Veränderungen des Oxidationszustandes des Cystein-Restes könnten die Konformation des RGD-Tripeptides verändern und somit die Affinität zu potentiellen Liganden beeinflussen (Bosse et al., 2000). Neben der RGD-abhängigen Wechselwirkung mit den Integrinen avb3 und a5b1 gibt es auch Hinweise, daß die Interaktion mit dem Integrin a9b1 RGD-unabhängig verläuft (Eto et al., 2000). Dabei wirkt sich die Anwesenheit von Magnesium(II)- und Mangan(II)ionen als stimulierend, die Anwesenheit von Calcium(II)ionen als inhibierend auf die Interaktion aus. Die Interaktion von Integrinen mit der extrazellulären Matrix zeigt die gleiche Sensitivität gegenüber den Konzentrationen von Magnesium(II)-, Mangan(II)- und Calcium(II)ionen (Chen et al., 1999a; Eto et al., 2000).
Die Cystein-reiche Domäne von ADAM 15 ist hinsichtlich ihres Einflusses auf die physiologische und pathologische Funktion des Moleküls noch nicht charakterisiert worden. Ihre Beteiligung an der korrekten Prozessierung der Proform in Analogie zu der Cystein-reichen Domäne von ADAM 17 wird diskutiert (Milla et al., 1999). Die Cystein-reiche Domäne von ADAM 15 weist zudem keine hydrophobe Region auf, so daß ihre Beteiligung an Zell-Zell-Fusionsprozessen in Analogie zu Meerschweinchen- oder Rinder-Fertilin a, ausgeschlossen wird (Muga et al., 1994; Krätzschmar et al., 1996; Waters und White, 1997).
Die zytoplasmatische Domäne enthält zwei Prolin-reiche Regionen PPPPRKP und RPAPPPP, so daß eine Wechselwirkung mit SH3-Domänen-tragenden Proteinen seit der Klonierung der cDNA in Betracht gezogen wurde (Krätzschmar et al., 1996). Somit könnte dieser Domäne eine Bedeutung bei der Weiterleitung intrazellulärer Signale oder eine Funktion bei der Anbindung an das Zytoskelett zukommen (Krätzschmar et al., 1996). Tatsächlich wurde für die Proform von hADAM 15 gezeigt, daß sie mit den beiden SH3-Domänen-tragenden Proteinen Endophilin I und SH3PX1 interagiert (Howard et al., 1999). Die reife Form von hADAM 15 (ohne Pro-Domäne) zeigte jedoch keine Interaktion mit Endophilin I und SH3PX1. Es wird vermutet, daß die Interaktion mit der Proform die subzelluläre Lokalisation oder die Funktion von hADAM 15 beeinflußt. Diese Interaktion müßte somit noch vor dem medialen- bzw. trans-Golgi-Apparat stattfinden, da hier die Pro-Domäne entfernt wird (Lum et al., 1998). Aus diesem Grund wird eine Beeinflussung der Zell-Zell-Interaktion durch SH3-Domänen-tragende Proteine ausgeschlossen, da hierfür die SH3-Domänen-tragenden Proteine mit der reifen Form von hADAM 15 interagieren müßten (Howard et al., 1999).
Die intrazelluläre Prozessierung von ADAM-Proteinen wurde erstmals am Beispiel des murinen ADAM 15 untersucht. Bei der Untersuchung von Mausgewebe konnte festgestellt werden, daß der Großteil des dort detektierten mADAM 15 ohne Pro-Domäne vorliegt (Lum et al., 1998). Überraschend war jedoch die Beobachtung, daß in mit mADAM 15 transfizierten COS-7 Zellen mehr als die Hälfte des prozessierten Proteins intrazellulär vorkommt. Möglicherweise befördert ein endozytotischer Vorgang das reife Protein nach der Integration in die Plasmamembran wieder in das Zellinnere zurück (Lum et al., 1998). Für ADAM 17 konnte ebenfalls gezeigt werden, daß es nach Prozessierung vorwiegend in einem intrazellulären Kompartiment lokalisiert ist (Schlöndorff et al., 2000). Ohne Erklärung blieb die Beobachtung, daß die Pro-Domäne bei auf der Zelloberfläche exprimierten mADAM 15 trotz proteolytischer Prozessierung der Furin-Spaltstelle immer noch mit dem Restprotein assoziiert bleibt und darüber hinaus an mehreren unbekannten Positionen eine proteolytische Spaltung erfährt (Lum et al., 1998).
Bisher ist die Expression von ADAM 15 mit zwei pathologischen Phänomenen korreliert worden. Einerseits konnten Herren et al. 1997 zeigen, daß das Auftreten arteriosklerotischer Läsionen mit einer verstärkten Expression von hADAM 15 einhergeht. Sie verwendeten zwei zum einen gegen die zytoplasmatische Domäne und zum anderen gegen die EGF-ähnliche Domäne gerichtete Antikörper zur immunhistochemischen Untersuchung von gesundem und pathologisch verändertem Aortagewebe von Primaten. Sie zeigten hierbei, daß nur im Fall einer arteriosklerotischen Läsion, eine Expression von ADAM 15 zu detektieren war. Neben diesen in vivo Untersuchungen wurde die Expression von ADAM 15 in humanen glatten Muskelzellen der Aorta und in humanen Endothelzellen der Nabelschnur in vitro nachgewiesen. Die Expression von ADAM 15 war konstitutiv und weder nach Aktivierung durch Zytokine oder chemoaktiver Reagenzien noch durch unterschiedliche Zelldichten zu beeinflussen. Ausgehend von den Ergebnissen der immunologischen Untersuchungen von humanen Endothelzellen der Nabelschnur wurde die folgende Möglichkeiten für die Prozessierung und die potentiellen Funktionen von hADAM 15 vorgeschlagen (Abb. 3).
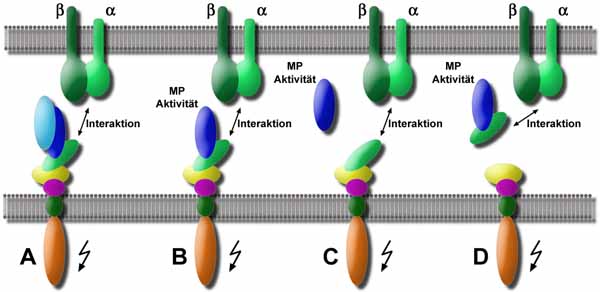
A) Die Disintegrin-Domäne könnte mit Integrinen anderer Zellen wechselwirken und so Zell-Zell-Kontakte und Signaltransduktionsprozesse initiieren. B) Durch die Freisetzung der Pro-Domäne kommt es zur Aktivierung der Metalloproteinase-Domäne, die in membrangebundener Form andere Zelloberflächenproteine der gleichen oder einer benachbarten Zellen proteolytisch prozessieren könnte. Darüber hinaus könnte die Disintegrin-Domäne mit Integrinen einer Nachbarzelle interagieren und zur Signaltransduktion beitragen. C) Ein weitere Möglichkeit besteht darin, daß die gesamte Metalloproteinase-Domäne zusammen mit der Pro-Domäne (hier nicht dargestellt) vom membrangebundenen Protein getrennt wird, in das umliegende Gewebe diffundiert und es dort zu einer Aktivierung der Metalloproteinase kommt. Die nun endständige Disintegrin-Domäne induziert dann möglicherweise Zell-Zell-Kontakte und Signaltransduktionsprozesse. D) Die letzte Alternative sieht die gemeinsame Abspaltung der Pro-, Metalloproteinase- und Disintegrin-Domäne vor, die möglicherweise durch ein intrazelluläres Signal initiiert wird. Durch die Disintegrin-Domäne zum Wirkort geführt, entfaltet die Metalloproteinase-Domäne nach Aktivierung dort ihr proteolytisches Potential (Herren et al., 1997).
Ein weiteres Indiz für die Beteiligung von hADAM 15 in pathologischen Prozessen erbrachten Böhm et al. (1999). Sie konnten durch in situ-Hybridisierungsexperimente zeigen, daß das Knorpelgewebe im Fall einer osteoarthritischen Erkrankung eine intensive Expression der hADAM 15-mRNA aufweist. Insbesondere in den Chondrozyten ist die hADAM 15-mRNA-Produktion im Vergleich zu gesundem Gewebe merklich erhöht, die calcifizierte Zone ist jedoch annähernd frei von hADAM 15-mRNA-Transkripten. Darüber hinaus fanden sie auch einen Anstieg der hADAM 15-mRNA in Chondrosarcomazellen in vitro. Die Autoren stellten daraufhin die Hypothese auf, daß möglicherweise die Interaktion von hADAM 15 mit dem Integrin a5b1, welches ebenfalls in Chondrozyten exprimiert wird, die Anbindung des Integrins an die extrazelluläre Matrix stören könnte. In früheren Arbeiten konnte gezeigt werden, daß eine Störung der Interaktion einer Zelle zur Matrix über das Integrin a5b1 zu transformierten Zellen führt (Plantefaber und Hynes, 1989). Alternativ könnte hADAM 15 durch proteolytische Prozesse an der Umstrukturierung der Knorpel-Matrix beteiligt sein (Böhm et al., 1999).
Bei Zusammenfassung aller bisherigen Erkenntnisse über ADAM 15 kristallisieren sich folgende Merkmale heraus: ADAM 15 zeigt sowohl eine RGD-abhängige als auch RGD-unabhängige Interaktion mit einer definierten Auswahl von Integrinen. Über die zytoplasmatische Domäne wird die Interaktion mit SH3-Domänen-tragenden Proteinen vermittelt. Dies spricht zum einen für eine Beteiligung von ADAM 15 an Signaltransduktionsprozessen oder auch für eine Anbindung von ADAM 15 an das Zytoskelett. Die Prozessierung von ADAM 15 findet im Golgi-Apparat statt, wobei Proprotein-Konvertasen, wie z.B. Furin, beteiligt sind. Nur ein geringer Anteil des in der Zelle exprimierten Proteins ist tatsächlich auf der Zelloberfläche präsent, der weitaus größere Teil befindet sich intrazellulär in einem perinukleären Kompartiment. Hinweise auf eine proteolytische Aktivität der Metalloproteinase-Domäne sind bisher nicht publiziert worden.