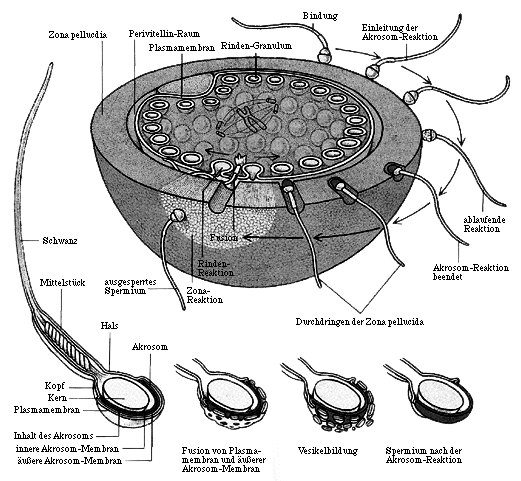
Die Fertilisation beim Säugetier ist das Ergebnis einer komplex abgestimmten Reihenfolge biochemischer Prozesse, die zur Bildung einer entwicklungsfähigen Zygote durch Fusion zweier Gameten mit haploidem Chromosomensatz führt. Die hierzu erforderliche Entwicklung der befruchtungsfähigen haploiden Gamete bei weiblichen Individuen erfolgt durch Oogenese der diploiden Urkeimzellen im Ovarium mit anschließender Follikelreifung. Bei der Bildung der männlichen Gameten wird zwischen der Spermatogenese in den Testis des männlichen Reproduktionstraktes und der Spermiogenese, also der Differenzierung zu reifen Spermien in der Epididymis unterschieden. Nach Ejakulation des Samens in den weiblichen Genitaltrakt erfolgt eine als Kapazitation bezeichnete Maturation der Spermatozoen, aus der die volle Befruchtungsfähigkeit der männlichen Gameten resultiert. Als Konsequenz dieser Vorgänge erwerben sie hyperaktive Beweglichkeit und die Fähigkeit zur Akrosom-Reaktion, die eine wichtige Voraussetzung für das Passieren der Zona pellucida (Glashaut) darstellt. Bei Säugetieren können mehrere Interaktionen zwischen Spermatozoen und Oozyte unterschieden werden. Zum einen binden Akrosom-intakte Spermien an die Zona pellucida, wodurch die Akrosom-Reaktion ausgelöst wird. Bei dieser Reaktion verändert sich der vordere Teil des Spermienkopfes, so daß die Penetration der Zona pellucida ermöglicht wird. Die Akrosom-reagierten Spermien penetrieren im Anschluß die Zona pellucida und binden an die Plasmamembran der Oozyte. Nach Fusion der beiden Plasmamembranen von Spermatozoon und Oozyte erfolgt die Initiation der zygotischen Entwicklungsprozesse. Abbildung 1 gibt einen Überblick über die Abläufe bei der Fertilisation.
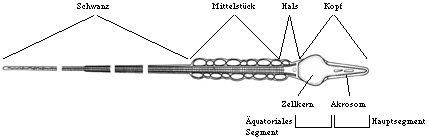
Spermien sind hochpolarisierte Zellen und weisen eine dreigliedrige Struktur, bestehend aus Kopf, Mittelstück und Schwanz, auf. Dieser Grundaufbau ist bei allen Säugetieren konserviert (McLeskey et al., 1998). Interaktionen mit der Oozyte finden allerdings nur im Bereich des Spermienkopfes statt, bei dem zwei Teilbereiche unterschieden werden können. Zum einen das Hauptsegment, das vor der Akrosom-Reaktion den vorderen Spermienkopf umfaßt, und das äquatoriale Segment, der verbleibende Rest des Spermienkopfes nach der Akrosom-Reaktion. Der vordere Teil des Spermienkopfes ist an der Bindung zur Zona pellucida beteiligt, während das äquatoriale Segment für die Einleitung der Gametenfusion verantwortlich ist.
Bei männlichen Individuen geschieht die Spermatogenese, also die Bildung der primären Spermatozyten, aus den Spermatogonien, welche über einen diploiden Chromosomensatz verfügen und aus den männlichen Urkeimzellen hervorgegangen sind. Die primären Spermatozyten werden durch die erste Reifeteilung in sekundäre Spermatozyten übergeführt, die nach einer zweiten Reifeteilung die Spermatiden mit haploidem Chromosomensatz ergeben. Die neu entstandenen Spermatozoen im Bereich der Testis sind infertil, und erst in der darauffolgenden Spermiogenese erlangen sie während der Passage durch die Epididymis die Fähigkeit zur Vorwärtsbewegung und die Fähigkeit, die Proteine der Zona pellucida zu erkennen und diese zu durchdringen. Bei diesem Spermien-Reifungsprozeß sind die Spermien und das epididymale Medium mitsamt des Epithels beteiligt (Perry et al., 1992), wobei viele der in der Epididymis wirkenden Proteine androgenreguliert sind. Ein wichtiger Aspekt des Reifungsprozesses ist die Tatsache, daß sämtliche Veränderungen der Spermien-Oberfläche nach Verlassen der Testis nicht auf der Neusynthese von Membranproteinen beruhen, sondern stets auf die Modifikation und Umstrukturierung bereits bestehender Membranproteine zurückzuführen sind (Myles und Primakoff, 1997).
Die Kapazitation beschreibt die physiologischen Vorgänge, die die Spermien im weiblichen Genitaltrakt erfahren. Nur kapazitierte Spermien sind in der Lage, die Cumulus-Schicht zu durchdringen (Saling, 1989) und die Eizelle im Anschluß zu befruchten. Die Cumulus-Schicht besteht aus den Follikelzellen des Ovars und einer von diesen sekretierten extrazellulären Matrix. Diese Schicht dient möglicherweise zur Bereitstellung einer adhäsiven Oberfläche, die das "Einfangen" der Spermatozoen erleichtern soll (Bedford, 1994; Camaioni et al., 1996; Eppig et al., 1996).
Eine zufriedenstellende Definition des Kapazitationsvorganges existiert bis dato noch nicht, doch lassen sich eine Reihe von Veränderungen während der Kapazitation beobachten (Tulsiani et al., 1997):
Die Bedeutung der Kapazitation für die vollständige Fertilität der Spermien wurde deutlich, als die erste in vitro Fertilisation von Hamster-Oozyten mit Spermatozoen gelang. Dazu mußten die Spermatozoen entweder vor der Imprägnation dem weiblichen Reproduktionstrakt entnommen oder aber epididymale Spermatozoen mit Eileiter-Sekreten vorinkubiert werden (Tulsiani et al., 1997). Diese Studien belegen, daß die Kapazitation von Faktoren ausgelöst wird, die im weiblichen Reproduktionstrakt angesiedelt sind. Bei den meisten Säugetier-Spezies kann der Ort der Kapazitation auf den Uterus und den Eileiter eingeschränkt werden. Die Spermatozoen erlangen ihre hyperaktive Beweglichkeit in der Isthmus tubä des Eileiters, wonach sie in die Ampulla einwandern, der Ort, an dem auch in den meisten Fällen die Fertilisation vonstatten geht (Tulsiani et al., 1997).
Die Akrosom-Reaktion ist ein irreversibler Vorgang, ähnlich der Exozytose somatischer Zellen. Dabei wird das Verschmelzen der Plasmamembran und der äußeren Akrosom-Membran (siehe auch Abbildung 1.1) durch einen intrazellulären Anstieg der Calciumkonzentration stimuliert, wodurch die Bildung von Hybridvesikeln initiiert wird (Snell, White, 1996). Diese werden zusammen mit dem Inhalt des Akrosoms in das umgebende Medium entlassen, wodurch weitere membranassoziierte, akrosomale Enzyme auf der nun exponierten inneren Akrosomen-Membran freigelegt werden. Der Calcium-Einstrom aus dem extrazellulären Raum wird dabei durch die Bindung des Spermatozoons an die Zona pellucida eingeleitet, wobei die Beteiligung von membrangebundenen Rezeptormolekülen diskutiert wird (Gong et al., 1995; McLeskey et al., 1998; Ward et al., 1994).
Die Zona pellucida ist eine mehr oder weniger stark ausgedehnte, durchscheinende extrazelluläre Matrix, die von der wachsenden Oozyte sekretiert wird und deren Plasmamembran sie umgibt (McLeskey et al., 1998). Sie paßt sich dabei dem Wachstum der sich entwickelnden Oozyte an, wobei ihre Ausdehnung (1 - 16 mm) charakteristisch für die betrachtete Spezies ist (Hinsch et al., 1997). Darüberhinaus schützt die Zona pellucida in den frühen Stadien der Entwicklung die Oozyte und den Embryo vor unerwünschten physiologischen Einflüssen.
Bei den meisten Säugetier-Spezies besteht die Zona pellucida aus drei sulfatierten Glykoproteinen (siehe Abbildung 1.3, linke Seite), genannt ZP1 (185-200 kDa), ZP2 (120-140 kDa) und ZP3 (83 kDa). Dabei bilden ZP2 und ZP3 eine filamentartige Matrix, die durch die ZP1-Moleküle quervernetzt werden (Hinsch et al., 1997). ZP3 ist für die Bindung der Spermien verantwortlich und maßgeblich an der Initiation der Akrosom-Reaktion beteiligt. Sein prinzipieller Aufbau ist in Abbildung 1.3 (rechte Seite) dargestellt.
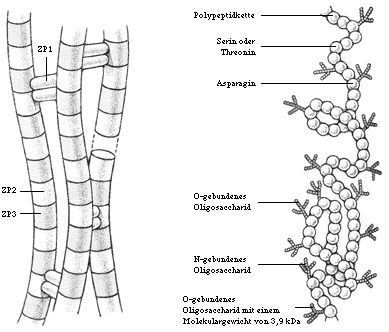
Die O-gebundenen Oligosaccharide mit einem Molekulargewicht von 3,9 kDa werden im wesentlichen für die Bindung der Spermatozoen an die Zona pellucida verantwortlich gemacht (Tulsiani et al., 1997).
Bei der Interaktion von Spermatozoen und Oozyte wird zwischen primärer und sekundärer Bindung an die Zona pellucida unterschieden. Die primäre Bindung der Akrosom-intakten Spermatozoen erfolgt an ZP3. An der sekundären Bindung der Akrosom-reagierten Spermatozoen ist ZP1 maßgeblich beteiligt. Darüber hinaus erfüllt es eine Funktion bei der Abwendung der Polyspermie (Myles und Primakoff, 1997; Töpfer-Petersen et al., 1997). Das ZP2-Molekül erfüllt vermutlich eine strukturstabilisierende Aufgabe. Die Spermien-Rezeptoren einer Eizelle erkennen und binden nur Samenzellen derselben Art, alle anderen werden verworfen (Wassarman, 1989). Eine der wenigen bemerkenswerten Ausnahmen sind Maultier- oder Mauleselembryonen, die aus der Vereinigung von Pferd und Esel hervorgehen. Die Nachkommen sind dann jedoch infertil.
Die Fusion von Spermatozoon und Oozyte ist mechanistisch erst wenig verstanden. Es ist jedoch bekannt, daß eines der durch die Zona pellucida vorrückenden Spermien den Pervitellinraum, einem schmalen Hohlraum zwischen der Zona pellucida und der Plasmamembran der Eizelle, erreicht, wonach es zur Verschmelzung der Plasmamembranen von Spermatozoon und Oozyte kommt (siehe Abbildung 1.1). Durch die Fusion wird eine Folge von Reaktionen ausgelöst, die binnen Sekunden die Plasmamembran verändert und binnen Minuten, in der sogenannten Zona-Reaktion, auch die Zona pellucida. Durch diesen Vorgang werden sowohl die Plasmamembran als auch die Zona pellucida für weitere Spermien undurchdringlich, so daß auf diese Weise eine tödlich endende Polyspermie verhindert wird (Wassarman, 1989). Divalente Kationen sind für eine erfolgreiche Gametenfusion essentiell, insbesondere Calcium kommt eine entscheidende Rolle bei den Adhäsions- und Fusions-Vorgängen zu (Evans et al., 1995). Eine Beteiligung membrangebundener Proteine auf Seiten der Spermatozoen und der Oozyte wird diskutiert, wobei die Aufklärung ihres Wirkungsmechanismus Gegenstand der aktuellen Forschung ist (Snell und White, 1996).
Eines dieser Proteine - das Fertilin - gehört zu einer Klasse von erst Ende der 80er Jahre entdeckten membrangebundenen Proteinen, die bei vielen Zell-Zell-Interaktionen eine entscheidende Rolle zu spielen scheinen. Diese als ADAMs (a disintegrin and metalloproteinases) bezeichneten Proteine zeigen große strukturelle Homologien zu einigen der löslichen Protein-Bestandteilen (SVMPs), die in Schlangengiften gefunden werden.